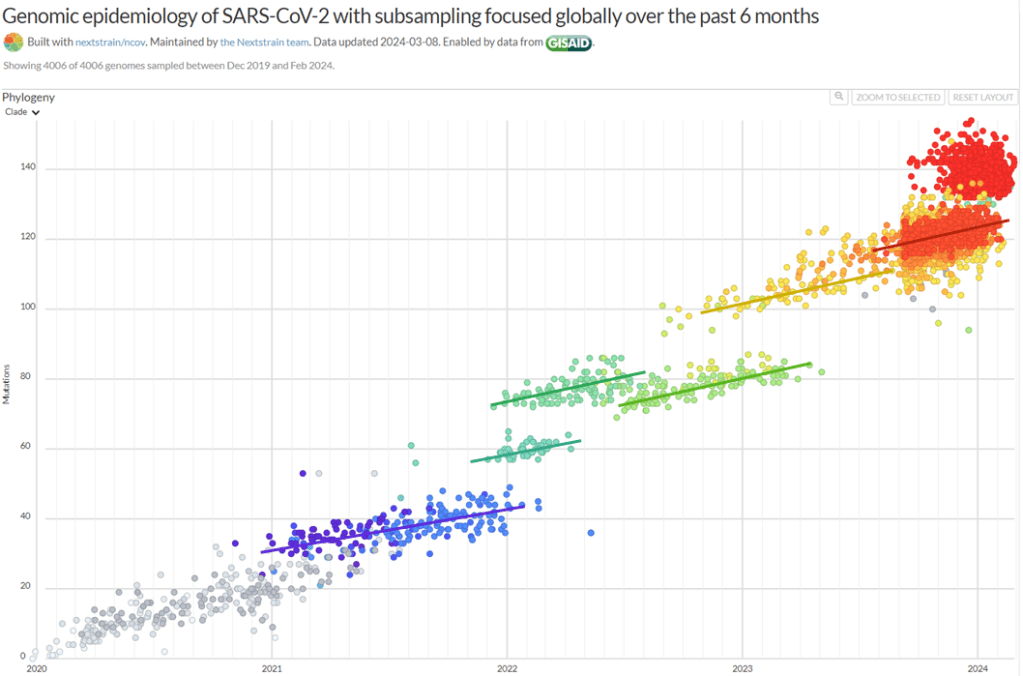
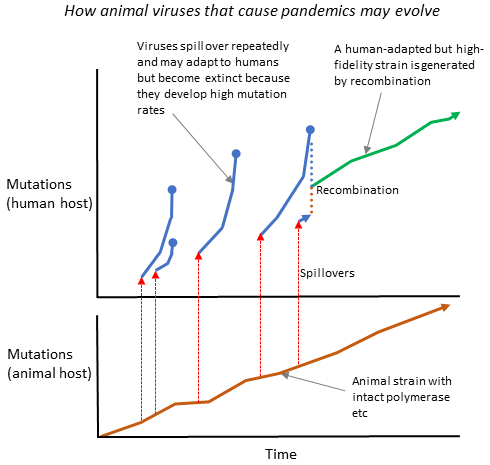
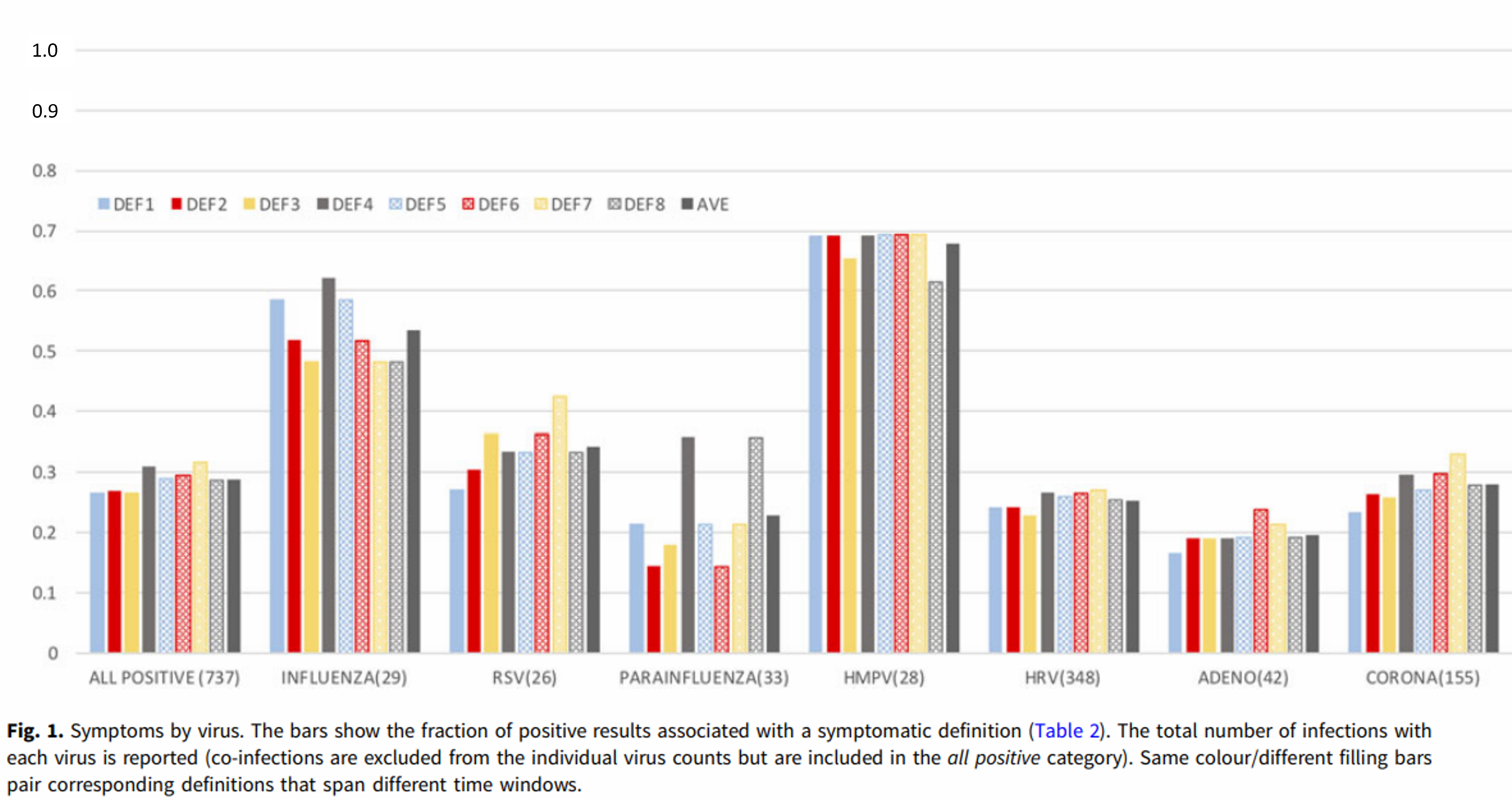
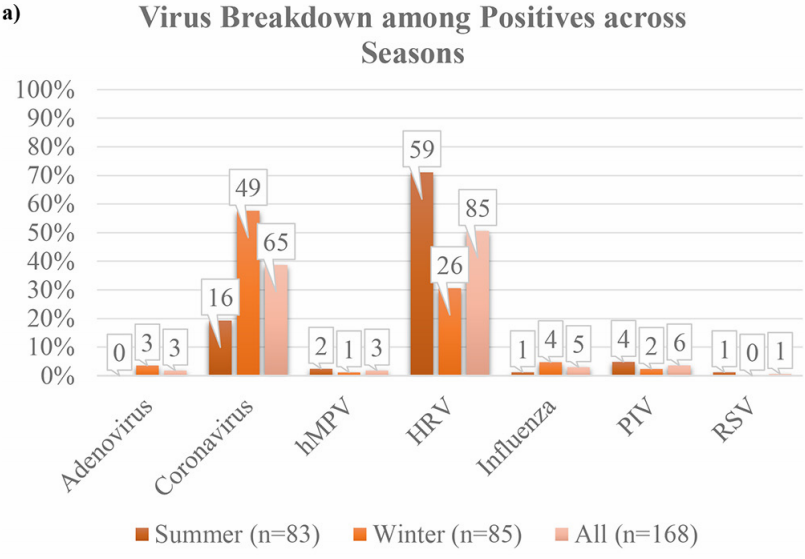
For detailed scientific information about respiratory viruses, including discussion of the trade-off model, viral dormancy and much else, see my 2021 paper:
Also
_________________________________________
Video introducing the hypothesis and describing the experiments that we need
Video that goes into the theory of TDVT
The TDVT Hypothesis
The temperature-dependent viral tropism (TDVT) hypothesis for respiratory viruses, referred to in my review as M4, is very simple:
-
-
- Viruses refrain from multiplying as fast as they can in order to spread themselves more widely (scientists call this the “trade-off” model)
- Respiratory viruses do this in a very simple way: they sense temperature. They refrain from developing at normal body temperature, and this allows them to keep out of the lungs, heart, brain etc. They normally replicate only in the cooler nose and throat, so the host keeps moving around and spreading the virus
- The seasonal pattern of viruses is a consequence of this. As temperatures rise in the air and thus in the nose and throat, the virus tends not to replicate in any part of the body.
-
Frequently Asked Questions
The strange arrivals – and departures – of influenza epidemics in the UK, 1946-1974
- The 8 major flu epidemics in the UK from 1946 – 1974 arrived very rapidly, then ceased within 3 – 5 weeks.
- Many susceptible individuals remained in the population, so the rapid ending of the epidemics is unexplained.
- Around 50% of influenza cases in New York City were recently shown** to be asymptomatic.
- It seems likely that new flu strains arrive in the community unnoticed, and are activated by temperature drops.
Applications to Covid-19
Predicting the seasonality of Covid-19
-
-
- Seasonality of respiratory illness is not mainly driven by changes in the survival rate of viruses outside the body. If it were, respiratory illness would be rarer in the Tropics than in the summer in temperate regions.
- We need an explanation of seasonality that fits all the facts.
- Seasonality seems to be a side-effect the natural temperature-sensitivity of virtually all respiratory viruses
- CoV-2 may be less temperature-sensitive than typical respiratory viruses, therefore less seasonal
- Experiments are needed!
-
Epidemiology of respiratory illness
-
-
- Respiratory illness responds so quickly to changes in ambient temperature that conventional epidemiology can’t give a good description
- Data often show a “harvesting” mechanism, where temperature changes activate dormant colds and flu
- Colds and flu frequently appear in the rainy season in the Tropics
- Explanations based on increased transmission in cold, dry places can’t work in the Tropics!
-
Covid 19 and the trade-off model
-
-
- Evolution balances the benefits of virulence in parasites (greater rate of shedding) against the disadvantages (shedding for less time, and less dispersal)
- “Short-sighted selection” increases virulence. “Far-sighted selection” increases transmission.
- Viruses that jump to new hosts are sometimes extraordinarily virulent. Well-established viruses clearly moderate their virulence.
- The benefits to the virus of moderation will increase as we approach herd immunity.
- Milder CoV-2 strains that are more temperature-sensitive may arise spontaneously in the next few weeks or months.
-
A simple model of CoV-2 transmission
-
-
- In institutions where there are many opportunities for transmission more virulent strains are favored
- Milder strains are favored in the community
- Lockdown slightly increases the competitive advantage of milder strains
- We should be very careful to contain virulent strains that may arise in care homes and other institutions where transmission can occur frequently
-
The adaptability of respiratory viruses
-
-
- Influenza tends to move from hotter countries to colder ones
- It seems to take a few months to adapt to a new climate
- CoV-2 replicated independently in the throat and lungs of a single patient
- The strain in the throat was derived from the lung one
- These observations give an indication of how rapidly respiratory viruses can adapt to new environments
- Experiments are needed ! !
-
Technical notes on CoV-2 for scientists
-
-
- We should focus on the temperature-sensitivity of RNA secondary structure
- The “s2m” structure in the 3’ untranslated region is of particular interest
- The distribution of ACE2 may not be the main driver of viral tropism
- Influenza has been shown to have a temperature-driven switch, with high temperature favoring transcription over replication
-
Suggestions for avoiding colds and flu – and Covid-19
-
-
- Observational studies can give us hints
- Never allow yourself to be chilled; dress warmly
- Take regular outdoor exercise, dressed warmly
- If you get a cold, stop exercise, avoid hot (and cold) drinks
- Stay warm!
-