Sexual complexity as a screen for replication fidelity: the Everest effect
My preprint can be found here:
Coughs, runny noses and fevers
We get more of them in the winter months. Could this be a side-effect of the natural temperature-sensitivity of almost all respiratory viruses?
Sexual complexity as a screen for replication fidelity: the Everest effect
My preprint can be found here:
March 2024
(Originally an X thread, dated 9 March 2024)
New thread – new theory
Is this unthinkable?
I think microbiologists have overlooked something huge—something that could have been understood at any time in the last 100 years if the right experiments had been done.
I start with the observation that the error rate when microbial genomic DNA/RNA is replicated must fluctuate due to natural variation.
Moreover, a demand for multiple mutations is created if a microbe is subjected to strong selective pressures.
The quickest way to deliver them may be to start by selecting mutations in the polymerase (those crosses and circles are supposed to be mutations!)

The idea is shown schematically below: consider a hypothetical virus with seven genes – a nucleic acid polymerase, three structural proteins, and three essential non-structural ones. Assume the virus has just spilled over to a new host, and beneficial mutations are required in all three structural proteins to adapt fully. Over time, sequential beneficial mutations may arise, as shown in Panel A, allowing the virus to adapt. If, however, a mutation first arises in the polymerase gene Panel B, a well-adapted strain may arrive much more quickly. However, such a strain would have a high mutation rate, and might be unstable because fatal mutations might arise too rapidly to be removed by “purifying” selection (Panel B).

A proposed lab experiment shown below encapsulates the same idea: under strong selection, microbes may become well-adapted (Series S1) but lose replicative fidelity (shown in Series S2).⬇️

But lineages with low fidelity have limited viability—because mutations will accumulate in essential genes (Series 2).
However . . . cultures containing these well-adapted but low-fidelity strains could, in principle, be “rescued” by recombination with high-fidelity strains – if they are available (see below and Series S3).

I think this happens constantly in nature and in the lab, but microbiologists don’t notice. It’s right in front of them, I suggest, whenever they “stress” their cells, or find one of their engineered constructs doesn’t replicate as strongly as they expected.
[Incidentally, IMO something similar happens in all complex organisms – which is why sexual reproduction is so popular with protists, fungi, plants, and animals – see https://vixra.org/abs/2303.0056 ]
What evidence supports this theory?
Well, first, there’s logic. If you demand a lot of mutations, fidelity is BOUND to fall by natural selection. But since life is still here, fidelity can be reinstated after a fall- which is where recombination, conjugation, etc, must come in.
Second, this proposal can explain why it’s sometimes difficult to propagate and maintain a stable strain of a newly-sampled bacterium or virus in the lab. For example, most archaea have not been isolated, and have been detected only from their gene sequences from environmental samples. Fidelity is often lost (I suggest) early on in the strong selective conditions of a lab, and recombination may not be feasible, making lab propagation difficult or impossible.
Then, there’s the extraordinary epidemiology of Covid-19. Why did the waves shown below ⬇️ arise so suddenly but then immediately collapse? They lacked the plateaus predicted by theory when immunity builds up slowly. I suggest these strains had slightly lower fidelity, allowing them to evolve reduced immunogenicity, etc., very rapidly but resulting in the subsequent accumulation of mutations in essential genes.


This can also explain how variants such as Delta, Omicron etc evolve. They may be the result of recombination between high- and low-fidelity strains.

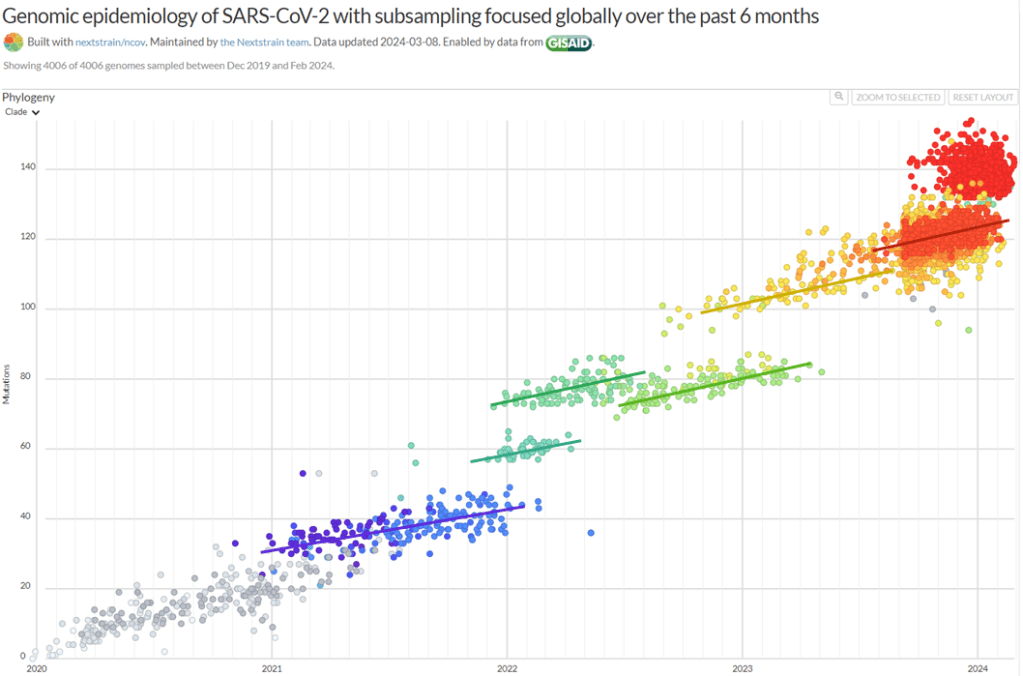
This theory can also explain why some SARS-CoV-2 variants have many mutations in Spike but much fewer in the rest of the genome and why an anomalously low proportion of the mutations in the spike of Omicron (and other variants) were C-to-T nucleotide “transitions”. In high-fidelity strains, most C-to-T transitions come from host modifications of viral RNA, whereas low-fidelity polymerases are expected to generate all nucleotide exchanges randomly.
Finally, we must explain why 100s of thousands of animal viruses can infect and often kill humans, but very rarely cause pandemics. Some, such as Lujo, Omsk, and Rift Valley viruses cause hemorrhagic fevers. Moreover, Marburg, Ebola, and Lassa have all been seen to spread from person to person. Lassa infects about 400,000 people, and kills around 5,000 every year (see Wikipedia entry), but, like the others, has never yet caused a pandemic. In the last 75 years, only seven pandemics have caused more than 10,000 deaths.
This is what I think happens: when an animal virus spills to humans, it rapidly adapts in the first patient and, in the process, develops a high mutation rate. For a pandemic to start, therefore, multiple spillovers need to occur until someone is infected simultaneously by well-adapted and high-fidelity strains, allowing recombination⬇️.
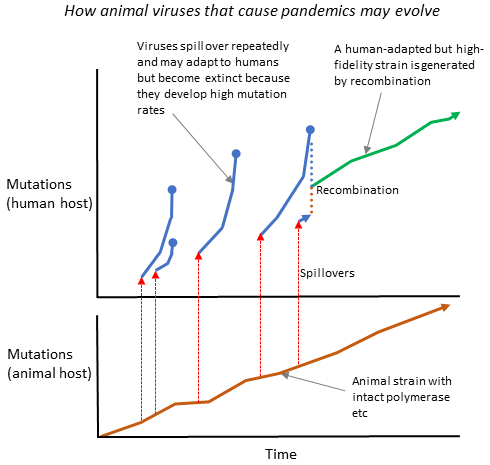
What happened to SARS-1? I guess it developed low fidelity, which recombination couldn’t restore because ongoing animal spillovers were prevented.
Best Origin of CoV-19 theory? SARS-2 was stable and well-adapted when it appeared, so repeated spillovers and recombination probably occurred over years in, e.g., South China. This may have resulted in a high-fidelity human-adapted virus. It seems likely scientists brought the virus to Wuhan, but it could have arrived by another route.
Was it “adjusted” in the lab along the lines suggested in the DEFUSE grant application? Or was a chimera constructed from different human or bat viruses? We don’t know.
So I’m not super-worried about future pandemics – as long as scientists don’t get involved, and as long as we don’t ignore repeated spillovers of animal pathogens in any particular region.
Does anyone know any papers—especially experimental papers —that can shed light on the conservation of replicative fidelity? Let me know.
What are the selective pressures acting on a respiratory virus?
When I wrote my scientific review of the seasonality of respiratory illness in Medical Hypotheses (2016) I devoted several paragraphs to trying to show that respiratory viruses could become dormant, lurking unseen in their human hosts without producing symptoms, and that they could be reactivated, often after changes in ambient temperature. I did this because at that time many scientists wouldn’t accept that the common respiratory viruses could possibly become dormant. In fact I was spurred on to write the article by a conversation that I had with a famous UK virologist; she told me that flu (as an example) was spread by an “endless chain of symptomatic individuals”. If I were to write that article now, I would cut out most of that section and just cite a paper that came out last year, describing the work done by scientists at Columbia University [1]. They tested residents of New York City for the presence of respiratory viruses throughout the year, whether or not they showed symptoms of a cold or flu. The results were remarkable. First, they found that roughly 60% of people carrying respiratory viruses were asymptomatic. Second, they found that people carried these viruses year-round: as many tested positive in summer as in winter.
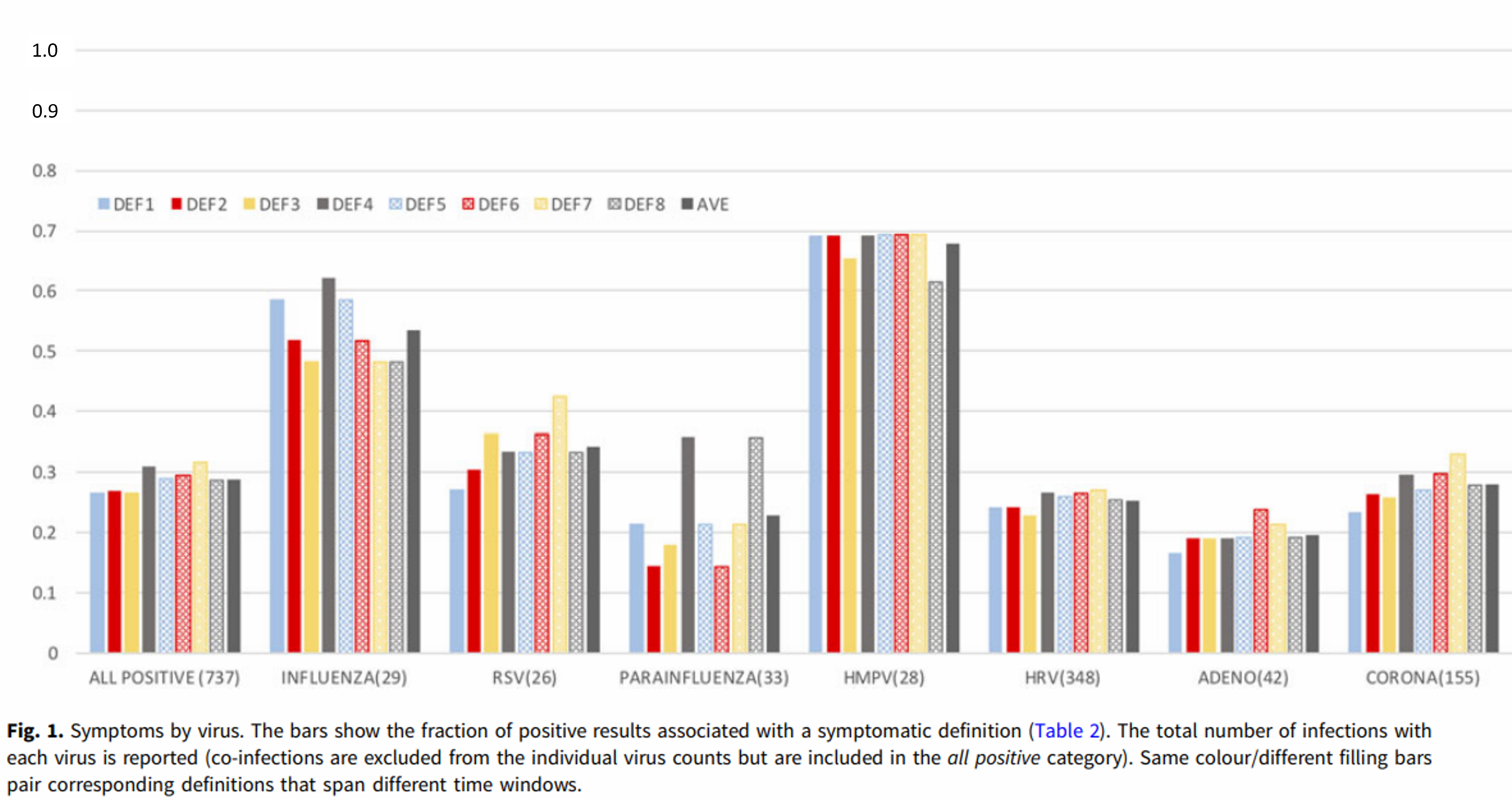
We’d like to make predictions of the likely course of epidemics such as Covid-19, so we’d like to understand the seasonality of the well-established human viruses. Therefore we need data about the seasonality of the presence of the virus. But we also want data about the seasonality of the illnesses caused. We’re going to want models, but they need to be more complicated, with a visible component – representing the people who are sick – and also a hidden component representing the asymptomatic cases where the virus is lurking undetected.
We will need to rethink our picture of respiratory illnesses in the light of the Columbia University studies. We can now see that respiratory infections are very largely asymptomatic and that respiratory viruses are, to a great extent, symbiotic. The body seems to get rid of them periodically, presumably when they get a little out of hand – often after chilling of the individual or cold weather. In any case, viruses were detected in only 17% of samples collected, suggesting that these infections come and go. Maybe the function of a cold is to have a clear-out of respiratory viruses and prevent more serious illnesses that might arise if the virus were to mutate and become virulent (it might, for example, lose some of its temperature-sensitivity).
For more thoughts on the Columbia University study and its implications for the mechanisms that underlie the seasonal appearance of respiratory illnesses click here.
Patrick Shaw Stewart, 9-14 May 2020
[1] Galanti, M., et al. “Rates of asymptomatic respiratory virus infection across age groups.” Epidemiology & Infection 147 (2019).
[2] Price, Rory Henry Macgregor, Catriona Graham, and Sandeep Ramalingam. “Association between viral seasonality and meteorological factors.” Scientific reports 9.1 (2019): 1-11.
Other papers on the NYC study:
Birger, Ruthie, et al. “Asymptomatic shedding of respiratory virus among an ambulatory population across seasons.” mSphere 3.4 (2018): e00249-18.
Galanti, Marta, et al. “Longitudinal active sampling for respiratory viral infections across age groups.” Influenza and Other Respiratory Viruses 13.3 (2019): 226-232.
_____________________
The TDVT Hypothesis, as explained by my friend Brian.
For a general discussion of the seasonality of respiratory viruses, written for the layperson, please see Every winter, colds and flu increase.
Shaw Stewart PD, Bach JL. Temperature dependent viral tropism: understanding viral seasonality and pathogenicity as applied to the avoidance and treatment of endemic viral respiratory illnesses. Reviews in Medical Virology. 2021 May 3:e2241. https://doi.org/10.1002/rmv.2241
I’ve just realized that I’ve been a bit slow on the uptake. I’ve been aware of the results of the surveys of colds and flu in New York City carried out by scientists at Columbia University [1-3] for a few days without spotting that they’ve supplied a particularly important bit of evidence in the puzzle of explaining, properly, the seasonality of colds and flu.
Obviously we get fewer colds and flu bouts in the summer than in winter, and this seasonality of respiratory tract viruses (respvirs) is a controversial topic, with many different explanations being put forward. Scientific reviews of the evidence, however, generally admit that there’s no good explanation for it [4, 5]. The trend is almost universal, though. With one known exception (parainfluenza type 3) every viral illness that is mainly transmitted by coughing, sneezing and runny noses is less common in temperate regions in the summer months than in the colder seasons. This includes completely unrelated viruses such as adenovirus (a DNA virus), influenza (an RNA virus that replicates in the cell nucleus), measles, mumps, RSV (three RNA viruses that replicate in the cytoplasm) and coronavirus (unusually, a “positive-sense” RNA virus). We can’t really claim to understand respvirs unless we can explain their seasonality.
Four broad mechanisms have been put forward by scientists to explain seasonality. The first two, which I labelled M1 and M2 in my 2016 review in Medical Hypotheses, are about transmission. M1 says we tend to crowd together more in cold weather, while M2 says that the virus can survive outside the body for longer in winter. What the Columbia University studies showed was that (1) around 70% of infections by respiratory viruses were asymptomatic , and, crucially, (2) that as many respvirs were present in the noses and throats of participants in the summer as in the winter. In fact, the most common respvir, rhinovirus, was more than twice as common in summer than winter! Overall, the investigators found that “across time, between 10% and 25% of the samples tested positive each week, but this overall rate of positivity did not exhibit a trend or seasonality” [3].
It’s taken me a few days to realize the importance of the second result. If the virus is present all-the-year-round, very often in an asymptomatic form, then we can rule out M1 and M2 as the main drivers of seasonality. We don’t need to look for reasons why these viruses can spread more in winter than in summer – if, in general, they don’t! (Of course I’m not saying M1 and M2 have no effect, just that they’re not the main drivers of seasonality.) So now we need to focus only on mechanisms that can convert asymptomatic infections into symptomatic ones.
That leaves the last two possible drivers, M3 and M4. M3 says that our immune defenses in the respiratory tract are weaker in winter than in summer. This is a much better candidate. Two American doctors, Mudd and Grant, showed in 1919 that when someone is chilled, for example by putting a wet towel on their back, the blood supply to their tonsils and pharynx decreases and the temperatures at these sites drop within a few minutes [6]. It’s not that our immune systems overall function worse in winter than summer: a study showed that vaccination in winter is, if anything, slightly more effective than vaccination in summer. But there seems to be a specific effect of chilling on our respiratory tract defenses.
However there are two problems with M3 as a seasonal driver of respvir illnesses: firstly, chilling didn’t show up as a cause of colds in the many classical studies that were carried out the 1950s and 60s. These studies normally used a “pedigree” cold-virus strain. That’s to say, the investigators took “snot” from someone with a cold, and put it into the noses of volunteers who were paid to have “holidays” at their centers and participate in experiments. (Cold viruses had not been well-classified at the time of many of these experiments, and the scientists sometimes had little idea of exactly what virus they were using; presumably they used trial-and-error to make sure that the sample they were using was safe!) Some of the volunteers were chilled, others kept warm. Which treatment they underwent, however, had no significant effect on the chance of getting a cold. This was repeated in many labs and over many years.
Now if M3 has such a powerful effect that it can drive the near-universal winter seasonality of so many respvirs, why didn’t it show up in these classical experiments?
The second reason that I’m skeptical of M3 – as a major seasonal driver – is that colds and flu are common year-round in the Tropics, much more common than in Europe and the USA at mid-summer. If our immune defenses work well during the summer here, why don’t they work in the Tropics where people are presumably less likely to be chilled?
That leaves M4, which I’ve proposed as the main driver of seasonality. I suggest that respvirs need a good way to stay in the nose and throat of their hosts, and to keep out of the lungs, heart, brain etc. If they go down into the lungs etc, they’re likely to make their host very sick, and – disastrously from their point of view – stop him or her from moving around and meeting other potential hosts. We usually stay at home and go to bed if we get fevers or muscle aches. Virtually all respvirs seem to have solved this problem in the same way: they’ve developed temperature-sensitivity. They replicate at temperatures below normal body-temperature, which normally means the nose and throat, and they become inactive at the higher temperatures of the lungs and other organs. Seasonality is, I suggest, a side-effect of this mechanism. As ambient temperatures increase in spring, the temperatures in the nose and throat also increase. The viruses then become less active, and we get fewer colds and bouts of flu. When fall arrives, and then winter, the temperatures in the nose and throat decrease as we breathe in the colder air. Now the virus becomes more active – so active that it may “accidentally” move down into the lower respiratory tract and cause fevers, possibly even getting into the blood-stream. This explanation, the TDVT hypothesis, is described in other posts in this blog.
Covid-19 has recently jumped into the human species from another animal. Therefore it’s not optimized to its host, and may not have much temperature-sensitivity when compared to well-established human respvirs. In practice, it seems to have retained some temperature-sensitivity because it often confines itself to the nose and throat, and also settles in the toes (causing chilblains). Like the nose and throat, the toes are some of the coldest parts of the body. Exactly how temperature-sensitive the virus is would be a very good subject for some urgent lab-work.
Patrick Shaw Stewart, 8 May 2020.
For more thoughts on the Columbia University study click here.
[1] Galanti, M., et al. “Rates of asymptomatic respiratory virus infection across age groups.” Epidemiology & Infection 147 (2019).
[2] Birger, Ruthie, et al. “Asymptomatic shedding of respiratory virus among an ambulatory population across seasons.” mSphere 3.4 (2018): e00249-18.
[3] Galanti, Marta, et al. “Longitudinal active sampling for respiratory viral infections across age groups.” Influenza and Other Respiratory Viruses 13.3 (2019): 226-232.
[4] Tamerius, James, et al. “Global influenza seasonality: reconciling patterns across temperate and tropical regions.” Environmental health perspectives 119.4 (2011): 439-445.
[5] Dowell, Scott F., and Mei Shang Ho. “Seasonality of infectious diseases and severe acute respiratory syndrome–what we don’t know can hurt us.” The Lancet infectious diseases 4.11 (2004): 704-708.
[6] Mudd, S., and S. Grant. “An experimental study of a possible mechanism for the excitation of infections of the pharynx and tonsil.” Am J Physiol 49 (1919): 144-145.
_____________________
The TDVT Hypothesis, as explained by my friend Brian.
For a general discussion of the seasonality of respiratory viruses, written for the layperson, please see Every winter, colds and flu increase.
Shaw Stewart PD, Bach JL. Temperature dependent viral tropism: understanding viral seasonality and pathogenicity as applied to the avoidance and treatment of endemic viral respiratory illnesses. Reviews in Medical Virology. 2021 May 3:e2241. https://doi.org/10.1002/rmv.2241
For a general discussion of the seasonality of respiratory viruses, written for the layperson, please see
Every winter, colds and flu increase
For detailed scientific information about the seasonality of respiratory viruses, including discussion of the trade-off model, viral dormancy and much else, see my 2016 paper:
For a discussion of the strange timing and duration of influenza epidemics, please see
The strange arrivals – and departures – of influenza epidemics in the UK, 1946-1974
For information about the probable seasonality of Covid-19, and whether we can expect it to become rarer in the summer, or reappear in the fall, please see
Predicting the seasonality of Covid-19
For comments about the epidemiology of Covid and other respiratory illnesses, please see
Epidemiology of respiratory illness
For discussion of how the trade-off model can be applied to the Covid epidemic see
Covid 19 and the trade-off model
For a simple model of the transmission of viruses such as CoV-2, please see
A simple model of CoV-2 transmission
For comments about how quickly we can expect viruses to adapt to new environments, please see
Adaptability or respiratory viruses
For more detailed scientific points about CoV-2, see
Technical notes on CoV-2 for scientists
For practical tips on avoiding respiratory illness see
Suggestions for avoiding colds and flu – and Covid-19
For information about viral seasonality please see
Shaw Stewart, Patrick D. “Seasonality and selective trends in viral acute respiratory tract infections.” Medical hypotheses 86 (2016): 104-119.
(A PDF is available: https://goo.gl/WexNkA )
or visit
For more information about funding and suggested lines of research please send a message to pshawstewart -at- gmail.com, or use the contact form (top right of this screen).
I’m not a professional virologist, but . . . . research into the seasonality of respiratory illness could be tackled at many levels of biological organization. For example:
For detailed scientific information about respiratory viruses, including discussion of the trade-off model, viral dormancy and much else, see my 2021 paper:
Also
_________________________________________
The temperature-dependent viral tropism (TDVT) hypothesis for respiratory viruses, referred to in my review as M4, is very simple:

